短链脂肪酸:连接饮食、肠道菌群与宿主健康的关键信使
2026-05-15
膳食纤维曾长期被视为一种无足轻重的食物成分,仅仅起到促进肠道蠕动的作用。
现代研究表明:膳食纤维的健康价值主要通过肠道菌群的代谢活动来实现,其核心产物是一类被称为短链脂肪酸(SCFAs)的小分子化合物。
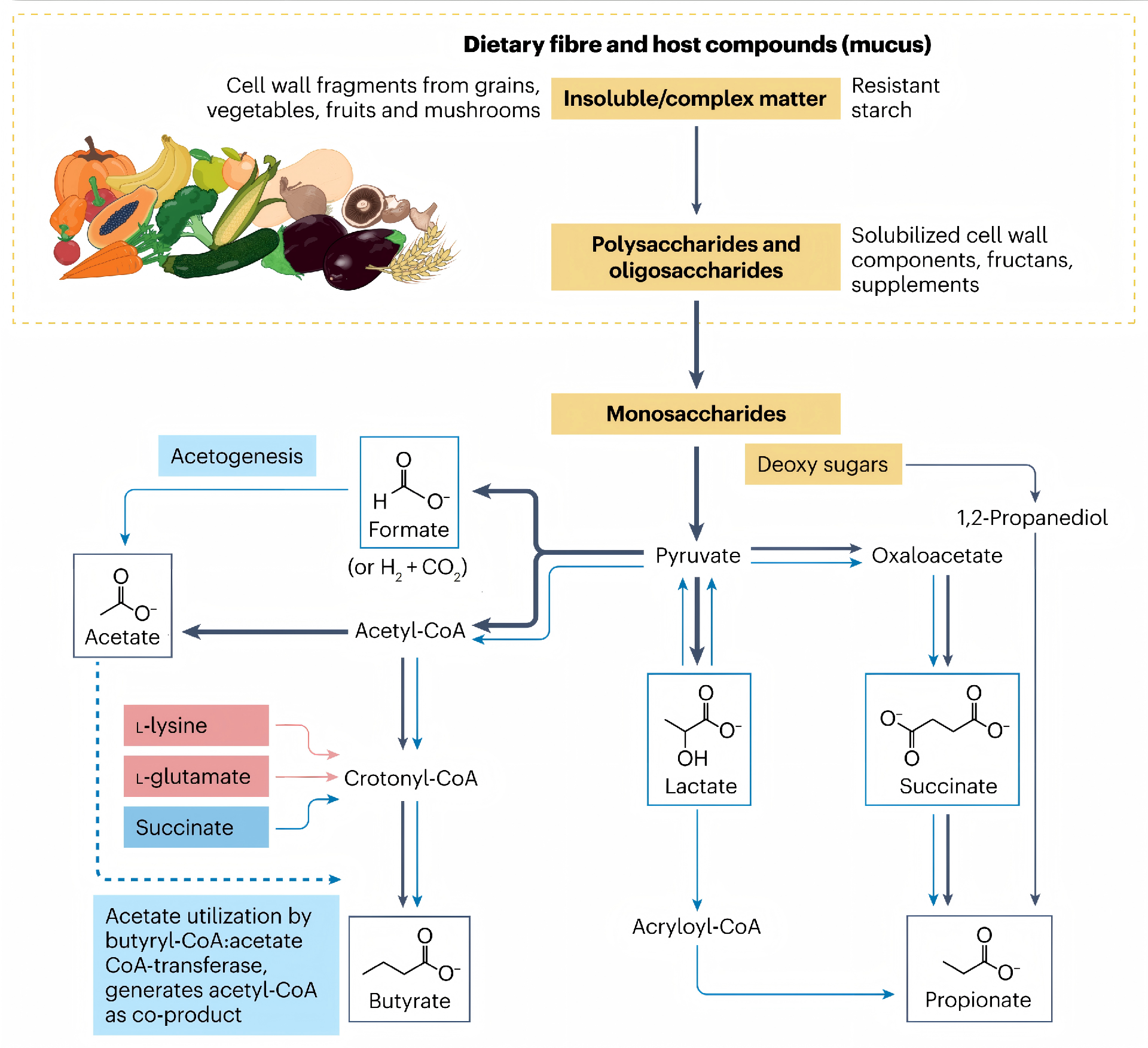
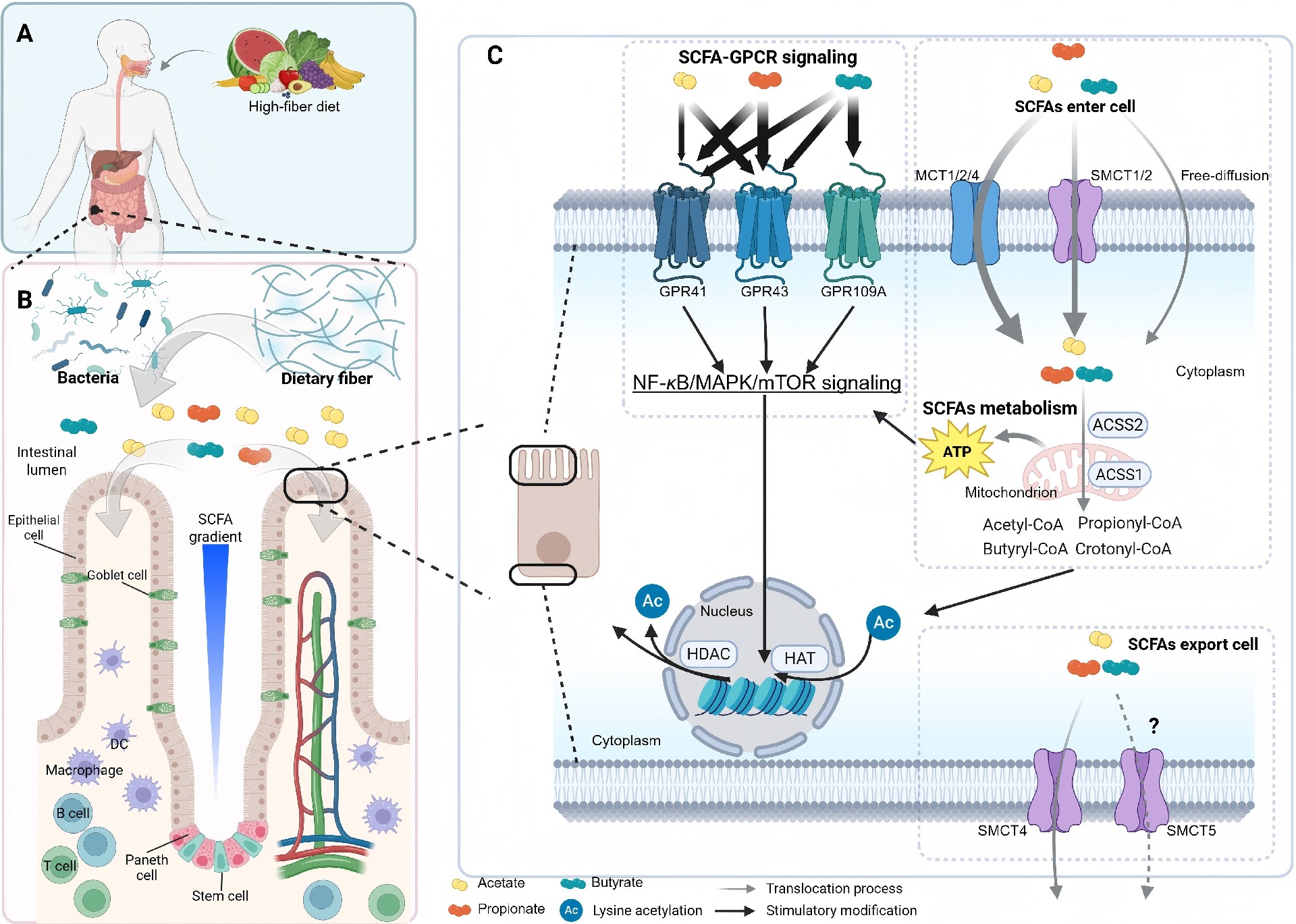
当不可消化的碳水化合物抵达结肠后,特定的厌氧菌群进行复杂的发酵: 将纤维转化为以乙酸、丙酸和丁酸为主的SCFAs。 这些分子并非简单的代谢废物,事实上,它们是连接饮食、菌群与宿主的关键信使。 超过90%的SCFAs会被肠上皮细胞吸收,其中: 丁酸 直接作为结肠上皮细胞的首要能量底物,为其提供约70%的能量需求; 丙酸 大部分经门静脉进入肝脏参与糖异生; 乙酸 相对更多地进入外周血液循环,抵达全身各组织器官。 这种差异化的代谢命运,使得SCFAs既能发挥局部效应,又能产生系统性影响。 近年来的研究进一步揭示:SCFAs的生理功能远超能量供应的范畴—— 它们通过激活细胞膜上的G蛋白偶联受体、抑制组蛋白去乙酰化酶以及参与细胞代谢重编程等分子机制,深度参与了肠道屏障维护、免疫调节、炎症控制乃至抗病毒防御等多个关键生理过程。 SCFAs的生成始于膳食纤维向结肠的完整递送。 这些源于植物细胞壁的多糖——包括纤维素、阿拉伯木聚糖、果胶以及抗性淀粉等——能够抵抗小肠内源酶的消化,最终完整地抵达结肠,为定植于此的数千亿微生物提供主要的碳源和能量。 结肠是一个高度厌氧的环境,绝大多数肠道细菌依赖发酵而非有氧呼吸来获取能量,这一代谢特征直接决定了SCFAs成为其发酵的必然终产物。 纤维的解构并非由单一菌株独立完成,而是依赖菌群成员之间精细的代谢分工与协作。 处于这一食物链顶端的被称为初级降解菌,它们配备丰富的碳水化合物活性酶,能够将复杂的多糖大分子切割为寡糖和单糖。这些水解产物不仅供自身利用,也会部分释放到肠腔环境中,为缺乏相应降解酶的次级发酵菌提供生长底物。 这种被称为交叉喂养的代谢互作模式,是维持菌群多样性和发酵效率的核心动力。 在酸性发酵产物中,乙酸、丙酸和丁酸占据主导地位,但三者的来源菌群谱系差异显著。 乙酸 生成途径最为广泛,绝大多数厌氧菌门类均可通过乙酰辅酶A途径产生乙酸; 丙酸 生产则相对集中,主要由拟杆菌门的成员通过琥珀酸途径完成,还有部分厚壁菌门的物种通过丙二醇途径利用脱氧糖生成丙酸。 丁酸 合成分布更为局限,几乎只限于厚壁菌门的特定类群,包括毛螺菌科的罗斯氏菌属、厌氧棒杆菌属以及瘤胃球菌科的柔嫩梭菌属等,这些细菌通过丁酰辅酶A转移酶途径缩合两个乙酰辅酶A分子生成丁酰辅酶A,再利用乙酸作为辅底物转化为丁酸。 肠道微环境同样深度调控SCFAs的产出谱系: 当发酵活跃导致肠腔pH下降时,许多丁酸产生菌较丙酸产生菌表现出更强的适应性,使得丁酸的相对比例随总SCFAs浓度升高而增加。 肠腔内氢气的积累也会促使某些细菌将代谢流从产氢转向产丁酸,以维持细胞内的氧化还原平衡。 环境条件与菌群组成的个体差异共同决定了每个人粪便中短链脂肪酸谱的独特性,这也为理解不同个体对同一膳食纤维干预的响应差异提供了理论基础。 图1. 短链脂肪酸生成的代谢途径 SCFAs进入宿主细胞后,主要通过三条相互交织的分子通路发挥其调控功能。 第一条通路涉及细胞膜表面的G蛋白偶联受体家族。 目前已知至少四种受体能够识别SCFAs: GPR43和GPR41分别对乙酸、丙酸和丁酸表现出差异化的亲和力; GPR109A被丁酸特异性激活; OLFR78可感受乙酸和丙酸。 这些受体广泛表达于肠上皮细胞、免疫细胞、脂肪细胞和神经元等多种细胞类型。 一旦配体结合,受体便激活与之偶联的异源三聚体G蛋白,引发下游信号级联反应: 包括抑制腺苷酸环化酶从而降低cAMP水平,或激活磷脂酶C产生IP3与DAG进而动员内质网钙库释放钙离子,最终通过MAPK等通路调控基因表达。 这一受体介导的快速响应机制,使SCFAs能够依据自身浓度梯度,在不同组织位置启动差异化的细胞应答。 第二条通路则深入到细胞核内部,通过表观遗传调控改变染色质结构和基因转录活性。 SCFAs中的丁酸和丙酸是I类和II类组蛋白去乙酰化酶的有效抑制剂。而组蛋白去乙酰化酶通常通过移除组蛋白尾部的乙酰基使染色质紧缩,抑制基因转录。 当SCFAs抑制其活性后,组蛋白乙酰化水平升高,染色质变得松弛开放,特定基因的表达随之增强。 这一机制在调节免疫细胞分化中尤为关键:例如丁酸通过抑制组蛋白去乙酰化酶促进叉头框蛋白P3基因座的高乙酰化状态,从而诱导调节性T细胞的分化和功能成熟。 丁酸的抑制效能最强,而乙酸的作用则相对较弱。 第三条通路涉及细胞能量代谢本身。 SCFAs进入线粒体后,乙酸和丁酸被转化为乙酰辅酶A,丙酸则转变为琥珀酰辅酶A,二者均汇入三羧酸循环驱动ATP生成。 对于结肠上皮细胞而言,丁酸是优先于葡萄糖的能量来源,其氧化供能约占其总能量需求的七成。 近年来针对免疫细胞的研究进一步揭示:乙酸被CD8阳性T细胞摄取后,通过乙酰辅酶A合成酶2转化为乙酰辅酶A,补给三羧酸循环代谢池,同时上调葡萄糖转运蛋白1的表达以增强糖酵解通量,最终提升干扰素γ和颗粒酶B的产生能力。 这种代谢重编程扩展了SCFAs从单纯供能分子到免疫效应增强子的功能内涵。 三条通路并非孤立运行,它们在信号网络和代谢节点上存在广泛交联,共同构成了SCFAs多层次调控宿主生理的分子基础。 图2. 短链脂肪酸的作用机制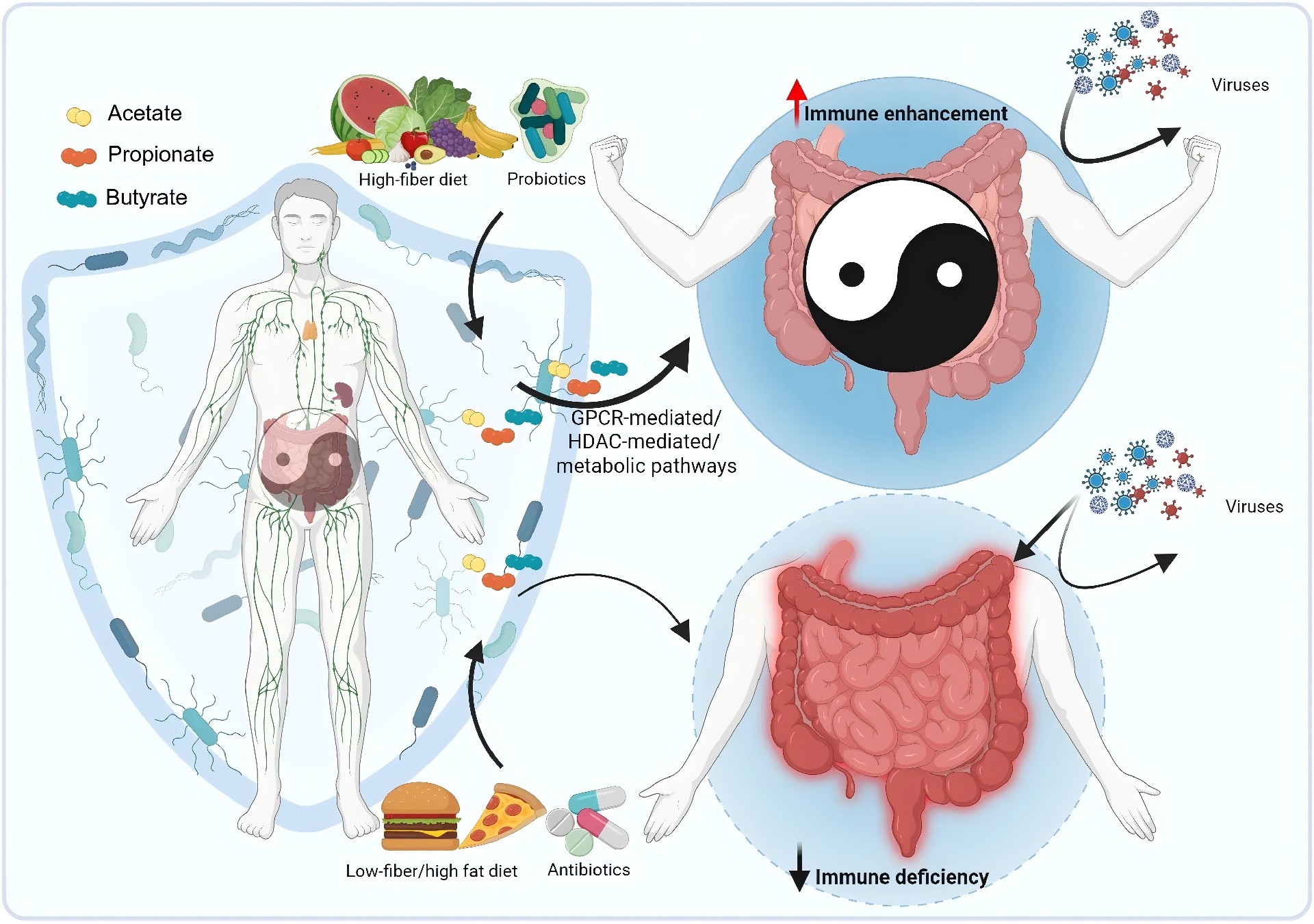
SCFAs在肠道局部浓度最高,因此其在肠道疾病中的作用最为直接和显著。
炎症性肠病(IBD)患者肠道中产SCFAs的细菌如普拉梭菌和肠罗斯氏菌显著减少,粪便可检测到的乙酸、丙酸和丁酸水平均下降,且丁酸氧化能力的受损使结肠上皮对丁酸的反应性降低。 这种“双重打击”——生产减少与利用障碍——共同加剧了肠道炎症。 临床试验显示,丁酸钠灌肠或口服微胶囊制剂可部分缓解溃疡性结肠炎和克罗恩病的症状,但疗效存在异质性,尚需更大样本验证。 在结直肠癌(CRC)中,丁酸呈现著名的“丁酸悖论”:正常结肠上皮细胞优先氧化丁酸供能,而癌细胞因瓦博格效应(Warburg effect)转向糖酵解,导致丁酸在核内积累,抑制组蛋白去乙酰化酶,进而诱导细胞周期阻滞和凋亡。 流行病学研究表明,高膳食纤维摄入与CRC风险降低25%相关,但粪便SCFAs水平与CRC的直接关联尚不明确,提示循环或组织中的SCFAs可能是更可靠的标志物。
SCFAs的作用远不止于肠道。在心血管与代谢疾病领域,高纤维饮食和SCFAs补充在动物模型中可降低血压、改善心功能。
但人临床试验结果不一:
口服丁酸对代谢综合征和2型糖尿病患者的血糖、血脂改善效果有限,却对儿童肥胖显示出体重指数降低的获益。 丙酸通过迷走神经反射增强餐后能量消耗和脂肪酸氧化,可能成为自主神经调节的潜在策略。 在呼吸系统疾病中,生命早期粪便中高水平的丁酸和丙酸与儿童哮喘风险降低显著相关,动物实验证实SCFAs通过GPR41依赖性途径减轻过敏性气道炎症。 神经系统疾病方面,阿尔茨海默病和帕金森病患者肠道菌群紊乱伴随粪便SCFAs减少,动物模型中丁酸和丙酸可保护血脑屏障、减少β-淀粉样蛋白沉积及抑制神经炎症。 多发性硬化患者血浆丙酸水平低下,口服丙酸可增强常规免疫治疗的效果。
这些发现共同勾勒出SCFAs从肠腔出发,经由循环系统调控远端器官免疫与代谢稳态的广阔健康版图。
图3. 短链脂肪酸在疾病中的作用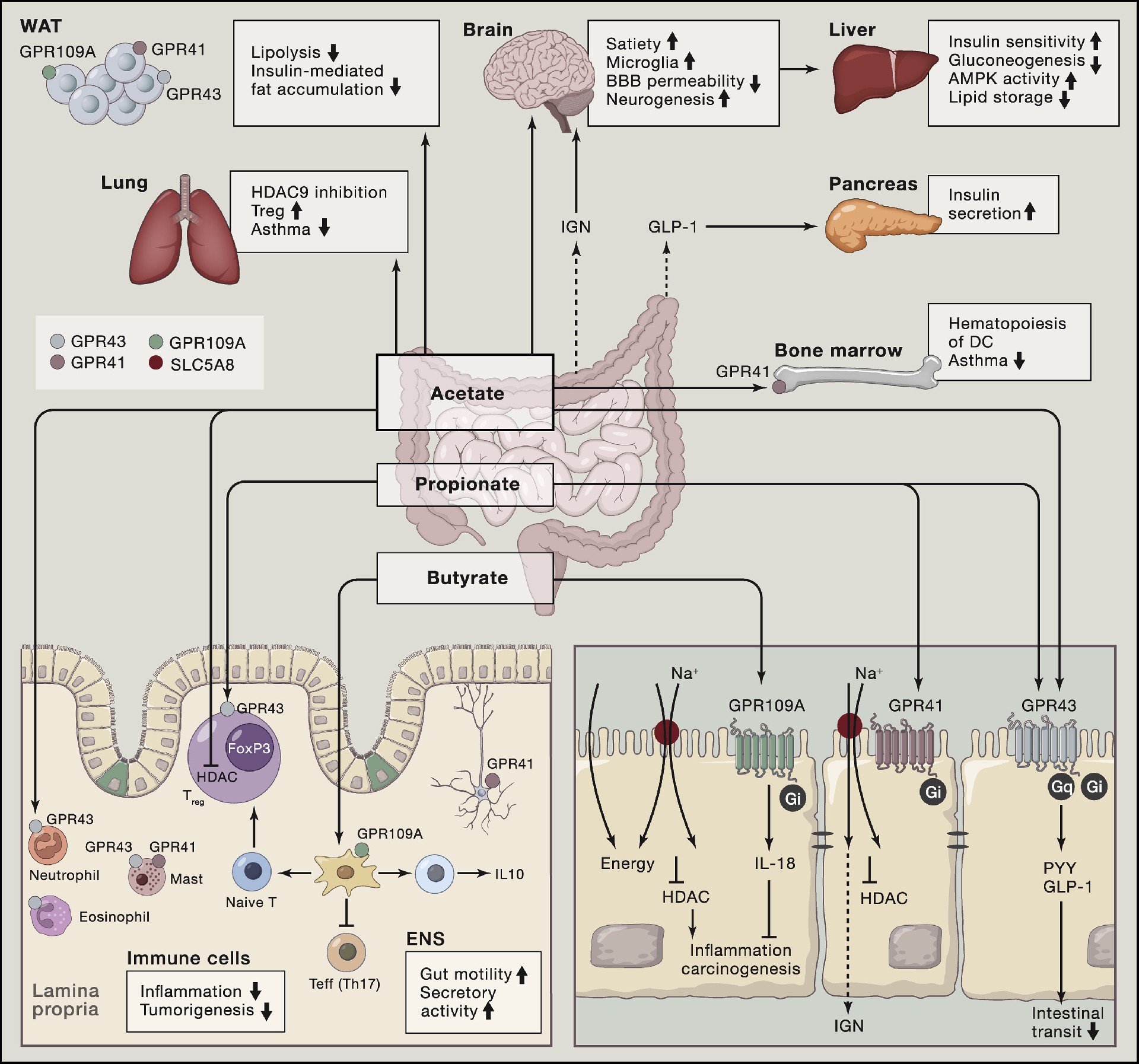
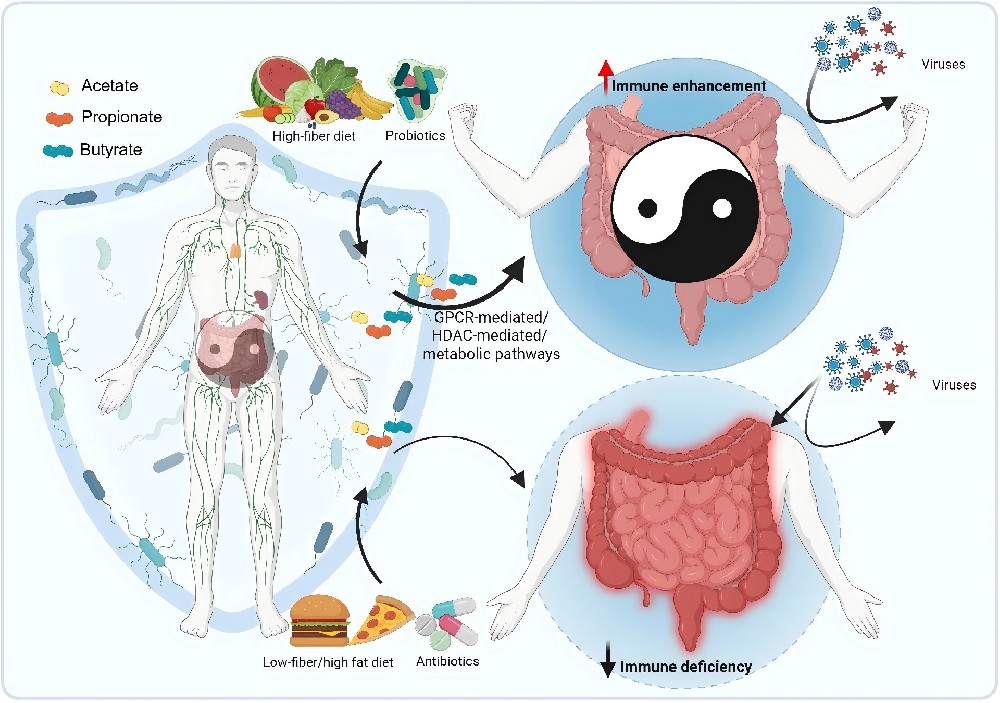
近年研究发现,肠道菌群产生的SCFAs不仅调控肠道稳态,更在宿主抗病毒防御中发挥关键作用。
SCFAs通过三条主要途径增强抗病毒免疫:
第一,激活G蛋白偶联受体GPR41和GPR43,进而启动MAPK信号级联,促进CD8阳性T细胞的代谢重编程与效应功能。 在流感病毒感染模型中,乙酸盐通过GPR43增强病毒特异性CD8阳性T细胞的干扰素-γ和颗粒酶B分泌,同时提高其线粒体氧化磷酸化和糖酵解水平,满足快速扩增的能量需求。 第二,丁酸盐和丙酸盐作为天然组蛋白去乙酰化酶抑制剂,可调节染色质可及性,上调干扰素刺激基因的表达。 然而,丁酸盐的作用具有细胞状态依赖性:在正常结肠上皮细胞中可抑制病毒复制,但在某些肿瘤细胞或高浓度条件下反而促进病毒复制,这与其抑制视黄酸诱导基因I介导的Ⅰ型干扰素通路有关。 第三,SCFAs还能维护肠道上皮屏障完整性,减少病毒经肠道向全身播散。呼吸道合胞病毒感染模型中,乙酸盐通过GPR43‑Ⅰ型干扰素受体轴增强肺上皮细胞的干扰素-β产生,降低病毒载量。
这些发现将SCFAs从传统的抗炎介质拓展为抗病毒免疫的新兴调节因子,为开发基于菌群代谢产物的抗病毒策略提供了理论依据。
图4. 短链脂肪酸的免疫调节作用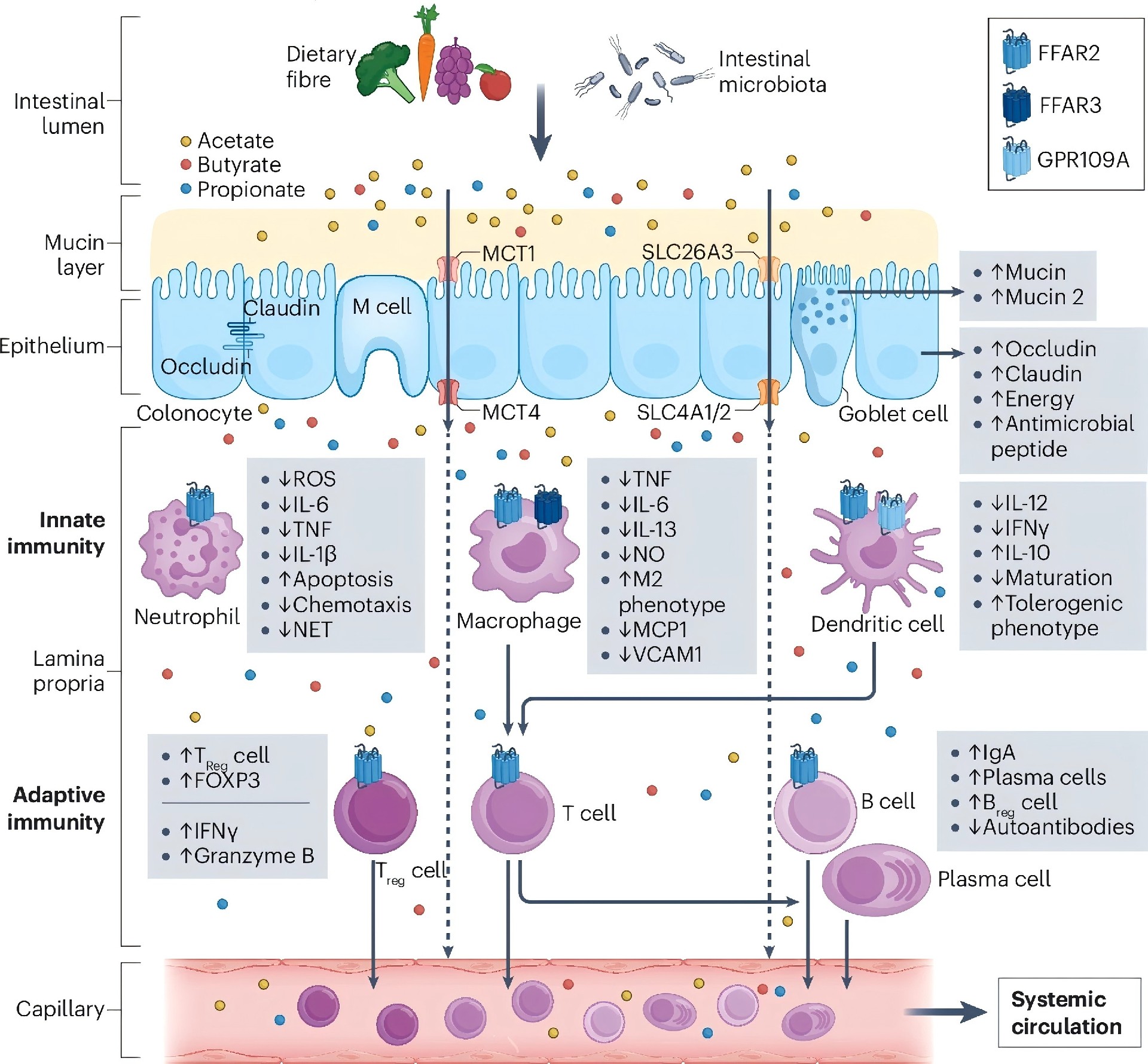
SCFAs的临床转化面临两大核心挑战:
递送效率与个体差异。
当前SCFAs可通过直肠灌肠、口服微胶囊或酯化淀粉等方式递送,其中微胶囊与肠溶包衣制剂能显著提高结肠靶向效率,但不同制剂在体内的释放动力学仍缺乏标准化评估。
膳食纤维干预作为提升内源性SCFA产量的策略,其效果受纤维类型、结构、剂量及个体菌群组成等多因素调控,单一纤维补充难以普适化。
机器学习与代谢网络建模逐渐被用于预测个体对膳食纤维的反应,为精准营养奠定基础。
此外,合成生物学手段正在改造共生菌株,使其携带特定SCFA合成基因,未来有望开发为“设计型”益生菌或菌群联合体。
粪便菌群移植虽可整体重塑菌群结构,但存在病原体传播风险,需谨慎评估。
总体而言,SCFA疗法的未来在于从“一刀切”走向个体化——依据患者菌群功能图谱、疾病类型和炎症状态,定制递送方案或益生菌组合。
同时,需进一步阐明SCFAs在肝、脑、肺等远隔器官的浓度-效应关系,以及长期补充的安全性边界。只有将基础代谢机制与临床转化工程学紧密结合,才能真正实现以SCFAs为靶点的精准医学。
1. Gut microbiota-derived short-chain fatty acids and their role in human health and disease. Nat Rev Microbiol. 2025;23(10):635-651. doi:10.1038/s41579-025-01183-w 2. Short-chain fatty acids: key antiviral mediators of gut microbiota. Front Immunol. 2025;16:1614879. Published 2025 Jul 25. doi:10.3389/fimmu.2025.1614879 3. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell. 2016;165(6):1332-1345. doi:10.1016/j.cell.2016.05.041 E N D
 技术咨询:
技术咨询: