多组学整合研究怎么做?两篇高分文章带你梳理研究思路
2026-07-14
近年来,多组学整合分析已成为生物医学研究的热门范式。从微生物组到宿主转录组,从代谢组到表观转录组,单一组学的“盲人摸象”时代正在被系统性的多层面解析所取代。那么,一篇优秀的多组学整合文章究竟该如何设计?研究思路又该如何搭建?
本文选取两篇近年发表的多组学研究,分别涉及肠易激综合征(IBS)、抗生素与表观转录调控,梳理其研究思路,供大家参考。
一.纵向多组学揭示肠易激综合征亚群特异性机制
文献标题:Longitudinal Multi-omics Reveals Subset-Specific Mechanisms Underlying Irritable Bowel Syndrome
发表期刊:Cell (IF=42.5,JCR Q1区,中科院1区)
发表时间:2020年9月10号
文章链接:https://doi.org/10.1016/j.cell.2020.08.007
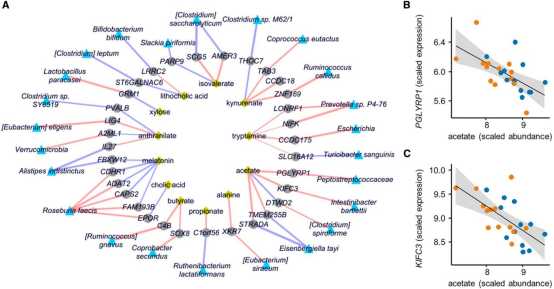
图1 Lasso正则化回归的多组学整合结果
研究问题:IBS(肠易激综合征)是一种高度异质性的功能性肠病,肠道菌群已被证实与多种慢性胃肠道疾病相关,但由于动物模型与人体研究存在脱节,且缺乏整合性的多组学视角,其机制性作用一直难以确定。
关键设计:同一个体在症状发作期和缓解期多次采样,形成自身对照。
组学设计:肠道微生物组(16S/宏基因组)、代谢组、宿主表观基因组和转录组。
组学 | 样本 | 技术平台 | 核心发现 |
微生物组 | 粪便+黏膜 | 鸟枪法宏基因组 + 16S | 菌群组成、功能基因通路(KO) |
代谢组 | 粪便+血清+黏膜 | 1H-NMR + LC-MS/MS + GC-MS | SCFA、色胺、胆汁酸、次黄嘌呤 |
转录组 | 结肠黏膜活检 | Bulk RNA-seq (TopHat→edgeR) | 差异表达基因、通路富集 |
甲基化组 | 结肠黏膜活检 | Illumina EPIC 850K 芯片 | 差异甲基化区域(DMR) |
核心思路:
· 1.纵向采样:对IBS患者进行多个时间点的样本采集,采用线性混合效应模型(LMM)或配对检验处理重复测量数据,消除个体间噪音
2.亚型分层:识别出IBS亚型特异性及症状相关的微生物组成和功能变异。
3.多组学分析:
(1)差异特征提取:分别在宏基因组(差异功能通路)、代谢组(差异代谢物)、转录组(差异基因)和甲基化组(差异甲基化位点)中筛选IBS相关的差异特征。(2)两两关联分析:
微生物组和代谢组关联:通过Maaslin2 相关性网络将特定菌属与特定代谢物关联,鉴定出显著代谢物-物种关联对;
功能基因和代谢物关联:分析基因组结构变异(DR/VR)与代谢物的相关性
代谢组和转录组关联:将微生物来源的代谢物与结肠黏膜的差异表达基因进行关联,筛选宿主响应通路。
转录组和表观遗传组关联:筛选出差异甲基化导致的差异基因
1. (3)多组学整合:通过Lasso回归等机器学习方法,将微生物组和代谢组数据与转录组和表观遗传差异整合,筛选出最有可能存在功能关联的“微生物-代谢物-宿主”变化子集。
4. 功能验证:(1)体外组织功能验证:使用人乙状结肠黏膜活检组织(42名参与者中采集),在 Ussing Chamber 中检测跨上皮离子分泌(短路电流 Isc),将多组学发现的代谢物变化(5-HT、色胺、初级胆汁酸)与宿主结肠生理功能直接挂钩。
(2)体外细菌培养验证微生物功能:基于 SV 关联分析选择菌株培养,验证多组学发现的"菌群降解次黄嘌呤"假说,LC-MS 检测培养上清液中次黄嘌呤水平
(3)无菌小鼠单定植:口服灌胃 ~2×10⁶ CFU 菌液;饮水中补充 100 μM 次黄嘌呤,定植后收集盲肠内容物,验证"特定菌群在体内确实能消耗次黄嘌呤"。
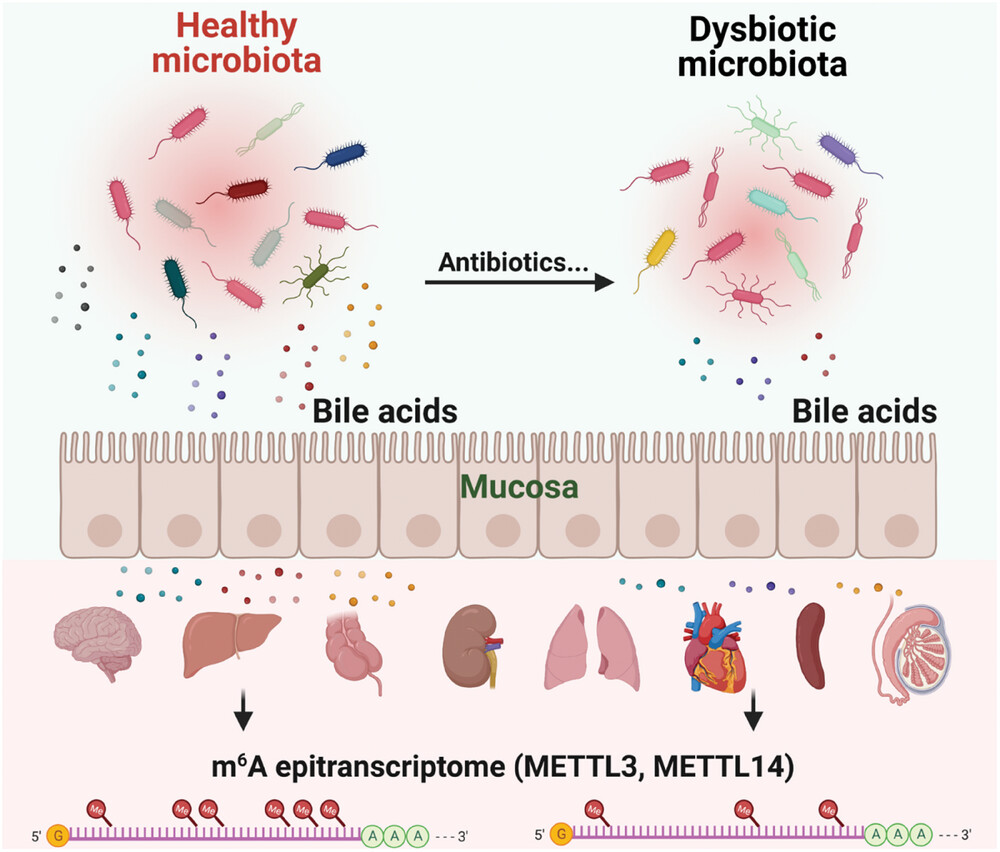
二.抗生素诱导的肠道菌群失调通过胆汁酸代谢调控宿主转录组和m6A表观转录组
文献标题:Antibiotic-Induced Gut Microbiota Dysbiosis Modulates Host Transcriptome and m6A Epitranscriptome via Bile Acid Metabolism
发表期刊:Advanced Science (IF=14.1,JCR Q1区,中科院1区)
发表时间:2024年5月7号
文章链接:https://doi.org/10.1002/advs.202307981
这篇发表在Advanced Science上的研究探索了肠道菌群如何通过代谢物重塑宿主的转录组和表观转录组。
研究问题:肠道菌群可通过代谢物影响宿主基因表达,但究竟是哪些代谢物、通过何种机制调控宿主的转录组和m6A表观转录组,此前尚不清楚。
研究假设:抗生素诱导的菌群失调 → 胆汁酸代谢紊乱 → 调控宿主 m6A 写入蛋白(METTL3/METTL14)表达 → 重塑多组织(脑、肝、肠等)m6A 表观转录组和转录组。
组学设计:采用多种抗生素诱导的菌群失调小鼠模型,结合粪便微生物移植(FMT) 到无菌小鼠,运用多组学方法进行研究。
组学 | 样本 | 技术平台 | 核心发现 |
微生物组 | 粪便(小鼠) | 16S rRNA 扩增子测序 (V4 区,Novaseq 6000) | 厚壁菌门/拟杆菌门比值失衡,变形菌门增加 产胆汁酸菌群丰度改变,菌群多样性显著降低 |
代谢组 | 粪便(小鼠) | 非靶向 UHPLC-MS/MS + 靶向胆汁酸 UHPLC-QTRAP MS(SCIEX 6500) | 胆汁酸代谢显著改变: 石胆酸、脱氧胆酸、7-酮基石胆酸下调; 牛磺胆酸上调;共鉴定 1725 种代谢物 |
转录组 | 脑、肝、肠、肾、肺 心、脾、睾丸(8 种组织) | Bulk RNA-seq | 组织特异性转录组重编程; 胆汁酸代谢和神经活性配体-受体互作通路在多组织中富集 |
表观转录组 | 脑、肝、肠/盲肠 | m6A-seq(MeRIP-seq,m6A-seq2 多重条形码方案,Novaseq 6000) | m6A 峰数量和分布显著改变; GGACU 基序富集; 神经信号和胆汁酸代谢通路相关基因 m6A 修饰改变 |
蛋白组 | 脑、肝、肠 | 无标记定量蛋白质组学(SDS-PAGE + LC-MS/MS, Q Exactive) | 差异蛋白富集于神经信号和代谢通路; m6A 写入蛋白(METTL3/METTL14)表达组织特异性改变 |
m6A 定量 | 脑、肝、盲肠(poly(A) 选择 RNA) | LC-MS/MS (Agilent 6490 Triple Quad, MRM 模式) | m6A/A 比值组织特异性改变(脑组织中显著降低); 验证 MeRIP-seq 结果 |
核心思路:
1. 模型构建:建立多种抗生素诱导的菌群失调小鼠模型,观察菌群变化对宿主的影响。
2. 多组学分析:
(1)16S rRNA 扩增子测序分析:抗生素处理组和无抗生素对照组相比,肠道菌群组成和丰度差异巨大。
(2)非靶向代谢组学检测:抗生素对肠道菌群的扰动改变了粪便样本中微生物来源的代谢物
(3)粪便菌群移植(FMT)到无菌(GF)小鼠,进行微生物组和代谢组分析,验证抗生素诱导的肠道菌群失调与胆汁酸代谢的关联,计算差异菌群和差异代谢物的相关性
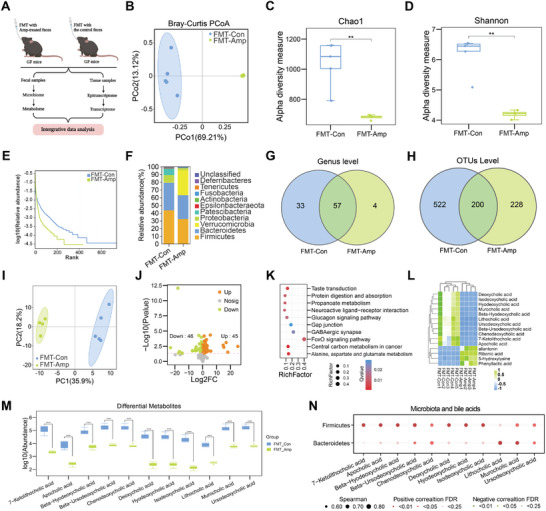
图2 在GF小鼠粪便微生物移植实验后进行的微生物组和代谢组分析
(4)八种小鼠组织转录组分析:筛选差异基因和富集通路,代谢-转录关联,重点关注胆汁酸代谢相关通路,使用定量蛋白质组学方法确定了菌群失调对宿主基因表达的蛋白质水平影响
(5)小鼠脑组织mRNA m6A 表观转录组分析:表明肠道菌群失调对小鼠脑 mRNA m6A 表观转录组具有全局性影响,KEGG 通路富集显示,肠道菌群失衡导致脑神经信号相关通路富集。
(6)肝和肠组织 m6A 甲基化组比较分析:m6A-转录相关性分析,m6A 修饰变化与基因表达变化的方向关系
(7)代谢-m6A 关联:研究m6A 峰变化与胆汁酸谱的关联,用多种胆汁酸(石胆酸、脱氧胆酸、7-酮基石胆酸、鹅去氧胆酸和猪去氧胆酸)处理三种哺乳动物细胞,以检验胆汁酸是否通过改变 m6A 写入蛋白直接影响宿主 m6A 表观转录组。
启示:这篇文章通过"抗生素诱导→FMT 因果验证→细胞机制验证"的三层递进设计,结合16S+代谢组+转录组+m6A-seq+蛋白组的五层组学数据,首次建立了"胆汁酸代谢→m6A 写入蛋白→表观转录组/转录组重塑"的完整分子链条,为菌群-宿主互作研究开辟了表观转录组学这一全新维度,每一层之间都有独立的实验验证(FMT、LC-MS/MS 定量、Western Blot、免疫荧光),而不是仅靠生物信息学拼接。这种"数据驱动发现 + 实验递进验证"的模式,是多组学整合研究从描述性走向机制性的标准范式。
总结:多组学整合研究的共性思路
梳理以上两篇文章,可以提炼出多组学整合研究的通用框架:寻找跨组学一致变化的生物学轴,并证明这条轴上的因果链条。
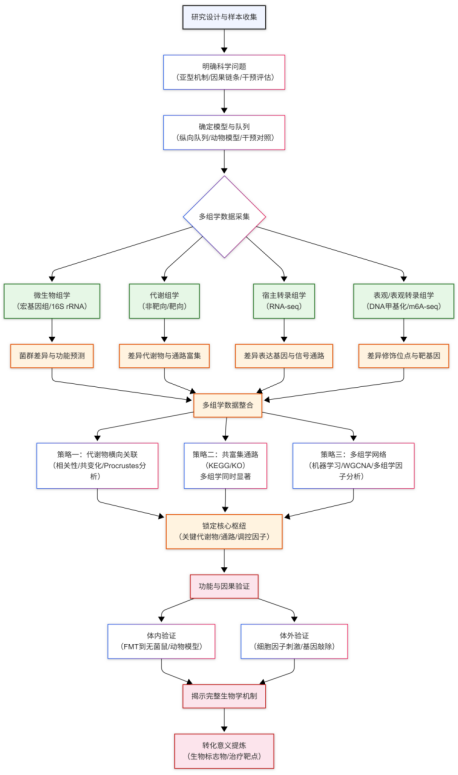
图3.多组学整合研究通用框架
第一步:明确科学问题
所有成功的多组学研究都始于一个清晰、可检验的科学问题。根据研究目标的不同,可以从三种基本范式中选择一种:
1.纵向追踪型:捕捉疾病随时间的动态变化,回答“什么变了”和“怎么变的”。适用于疾病进展、治疗响应轨迹等研究。
2.干预对照型:通过施加干预(药物、FMT、基因敲除等),回答“干预改变了什么”。适用于治疗机制、靶点评估等研究。
3.机制深挖型:在观察到现象的基础上,通过体外实验等手段,回答“如何改变的”。适用于从相关性走向因果性。
第二步:设计多组学策略
根据研究问题选择合适的组学层次。两篇研究的共同点是都包含了微生物组、代谢组和转录组,在此基础上可根据需求增加其他组学维度,如宿主遗传变异(基因组)、DNA甲基化/组蛋白修饰(表观组)、RNA修饰(表观转录组)等。
第三步:数据整合分析
1. 对每个组学数据进行独立分析,各自找出“有差异”的特征——差异菌属、差异代谢物、差异表达基因等。
2. 多组学整合分析:
机器学习整合 :面对“特征多、样本少”的困境,使用LASSO回归等算法从海量特征中筛选出对表型贡献最大的核心信号。适合在复杂数据中锁定关键分子或通路。
生物信息学整合 :使用Procrustes分析评估两个数据集整体结构的相似性,用O2PLS分离数据的“共有变化”和“特有变化”,用相关性分析构建跨组学分子网络。适合探索多组学间的协同变化模式。
实验验证整合 :用体外实验(如细胞因子处理)验证关键分子之间的因果关系,将“相关”升级为“因果”。适合在锁定关键分子后深挖调控机制。
3.通过整合分析,提炼出贯穿各组学的核心通路或调控轴
第四步:功能验证。
将整合分析发现的假说在独立体系中验证。体内验证(如FMT到无菌小鼠)可证实微生物群的致病性或治疗作用;体外验证(如特定代谢物处理细胞)可证实分子间的直接调控关系。最终落脚于生物标志物或治疗靶点的提炼。
第五步:临床/转化意义提炼
希望这份框架总结能为你的多组学研究设计提供参考。当然,具体实施时还需要根据自身的研究问题、样本资源和数据特点灵活调整,没有放之四海而皆准的模板,但有可遵循的思考路径。
 技术咨询:
技术咨询: